编者按:
随着对肠道菌群的不断研究,科学家们发现了越来越多肠道菌群与宿主代谢之间的关联,同时也越来越关注菌群与宿主代谢之间的因果关系,以及如何将因果关系产业化。而近年来的研究发现,菌群产生的代谢物或成这一因果关系的关键。
今天,我们特别对发表在 Molecular Cell 杂志上题为“From Association to Causality: the Role of the Gut Microbiota and Its Functional Products on Host Metabolism”的综述进行了解读,提炼了其中的重点与要点。希望本文为相关的产业人士和各位读者带来一些启发与帮助。
微生物的代谢产物
在成人肠道中,约含有 39 万亿个微生物细胞,与人体内的细胞数量相当甚至要更多一点,其中所包含的基因数更是人体基因的 1000 倍以上1,2。近年来,16S rRNA 和宏基因组测序技术的发展,使我们了解了微生物组成的变化与宿主代谢之间的联系,一系列的研究发现微生物组成会依据宿主饮食或使用的药物做出相应的改变3,6。
随着我们对肠道微生物了解不断加深,我们知道了:特定的肠道细菌与代谢疾病之间存在联系(表 1);肠道微生物的功能可能是菌株依赖的;饮食成分不仅能够影响微生物的组成还是许多微生物酶类的底物等等7~9。
而随着宏代谢组的发展,我们在体内发现了大量的微生物衍生代谢物,其中一些甚至是在宿主组织中发现的,这表明这些代谢物或可帮助宿主和微生物之间进行信息交流10。更进一步的研究表明微生物代谢产物对健康有着重要影响,因此,对微生物代谢产物与健康之间的因果关系和机制方面的研究变得越来越迫切11。
今天我们将从菌群移植、菌株、微生物代谢产物三方面简要介绍这三者与宿主疾病之间的因果关系与机制(图 1)。

图 1. 从关联性向因果性
菌群移植,初现因果
本世纪初,Jeffrey Gordon 实验室的一项关于肠道菌群的研究,打开了菌群移植的大门。该实验室的研究人员把肥胖小鼠盲肠来源的菌群转移到了无菌小鼠中,结果发现这些无菌小鼠也显示出肥胖的症状12,13。随后的研究进一步证实了粪菌移植(FMT)能够传递肥胖表型14。
此外,利用 FMT 治疗艰难梭菌感染(CDI)的过程中也出现了由于获得的是超重者个体的粪菌,导致患者变胖的例子15,16。
另一项研究,将从代谢健康的消瘦个体获得的粪菌移植到了代谢综合征患者体内,结果发现患者对胰岛素的敏感性增加17。
这些研究均表明肠道菌群与代谢性疾病之间存在因果关联:菌群移植会传递有益或者有害的代谢表型。但是利用 FMT 在治疗代谢性疾病具有一定的缺陷,比如治疗效果的维持时间有限,因此需要持续治疗,再者需要对捐献者进行严格地检查18。
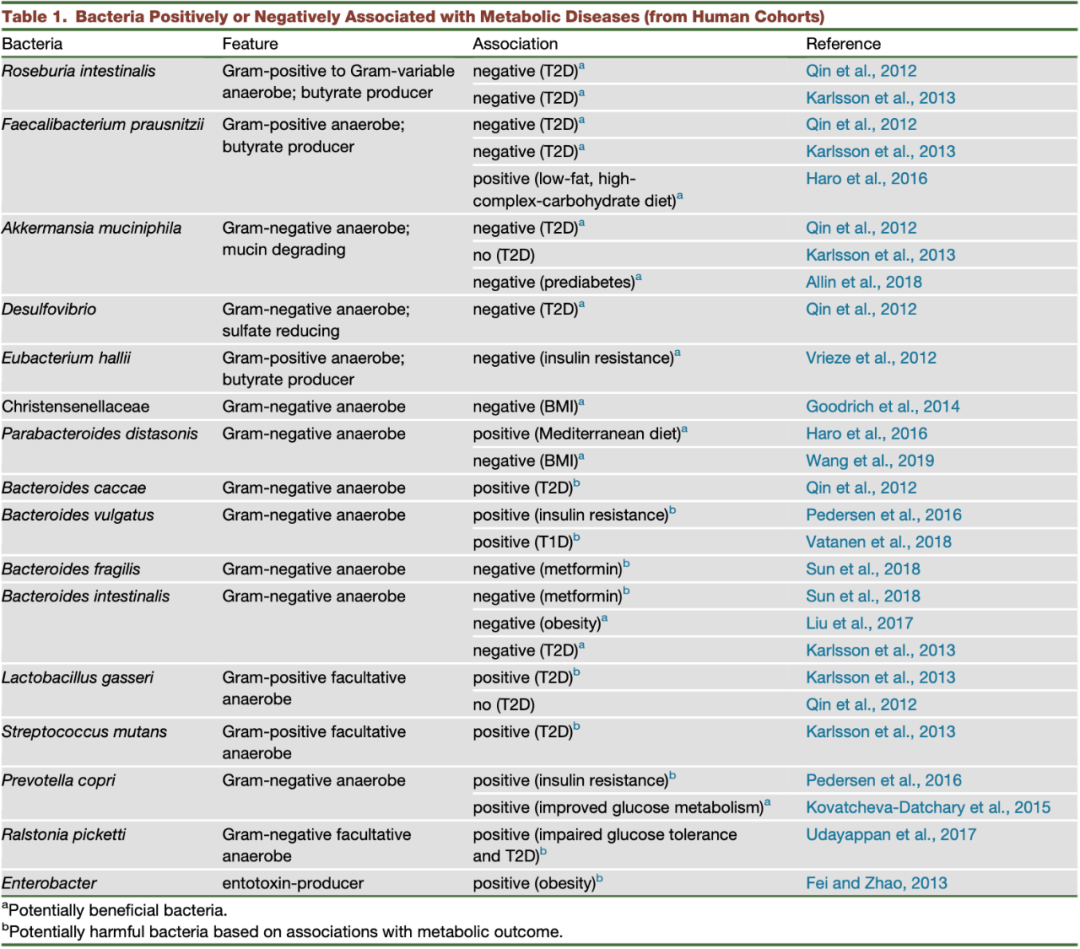
表 1. 与代谢疾病相关的阳性或阴性细菌(来自人体)
找到因果中的细菌
我们已经知道肠道微生物会影响宿主代谢,那么究竟是哪些肠道微生物在起作用呢?
Akk 菌
在人体肠道中有一种名为嗜粘蛋白艾克曼菌(Akkermansia muciniphila)的细菌,其相对含量较高,可以达到总量的 1%~4%(图 2),它可以对小鼠和人体的代谢起到一定的改善作用19:减少肥胖、葡萄糖不耐受和肠道渗透性20。
在发生胰岛素抵抗的人中,灭活的 Akk 菌能够在一定程度上改善高胰岛素血症和胰岛素敏感性,并且其上清液也具有一定的抗炎症作用21,22。灭活的 Akk 菌的益处可能来自于其外膜上的一种恒温蛋白,这种蛋白可以提高肠道屏障功能,并且对体重、脂肪葡萄糖代谢具有有益作用23。
普拉梭菌
在健康人体的肠道菌群中,普拉梭菌(Faecalibacterium prausnitzii)的占比超过 5%,它具有产丁酸的能力,还能分泌一种微生物抗炎症分子(MAM),对大肠炎小鼠具有保护效果24,25。
另外,研究人员发现这种菌与 2 型糖尿病之间存在负相关。但是由于其难以在小鼠中定植,所以目前还没有动物实验的数据证明其在葡萄糖代谢中的具体作用。此外,这种细菌产丁酸的能力具有菌株特异性且在不同宿主中的表现不同。而且其对氧极度敏感,所以难以培养保存。这些因素均限制了该细菌的研究与应用8,9,26。
霍氏真杆菌
霍氏真杆菌(Eubacterium hallii)是另一种产丁酸的细菌,可以提高胰岛素敏感性并增强糖尿病小鼠中的能量消耗,这表明它可能在疾病中具有保护作用17,27。
乳杆菌
肠道微生物中的乳杆菌(Lactobacillus)也具有改善代谢的作用。罗伊氏乳杆菌(L. reuteri)通过色氨酸衍生代谢物和配体激活的转录因子芳香烃受体(AhR)实现微生物-宿主信号交流,从而改善高脂饮食小鼠中不依赖体重的葡萄糖耐受性和胰岛素敏感性7。
其它细菌
还有其它细菌可能对宿主代谢存在积极作用。比如,给肥胖或者高脂肪饮食小鼠服用狄氏副拟杆菌(Parabacteroides distasonis)可能会通过提高微生物衍生物琥珀酸和次级胆汁酸的含量,抑制体重的增加,并预防高血糖症和肝脂肪变性28。
又如 Prevotella copri 或可改善葡萄糖代谢,因为其产生的琥珀酸和随后肠道糖异生相关29,30。另外,有研究表明 Parabacteroides goldsteinii 可以缓解肥胖、炎症和胰岛素抵抗,并且还能提高脂肪褐变标志的表达31。
虽然这些菌株对代谢是有效果的,但是这些效果可能具有菌株依赖性,还可能需要其它微生物的支持,并且存在个体差异,因此在人体中的作用需要进一步验证。
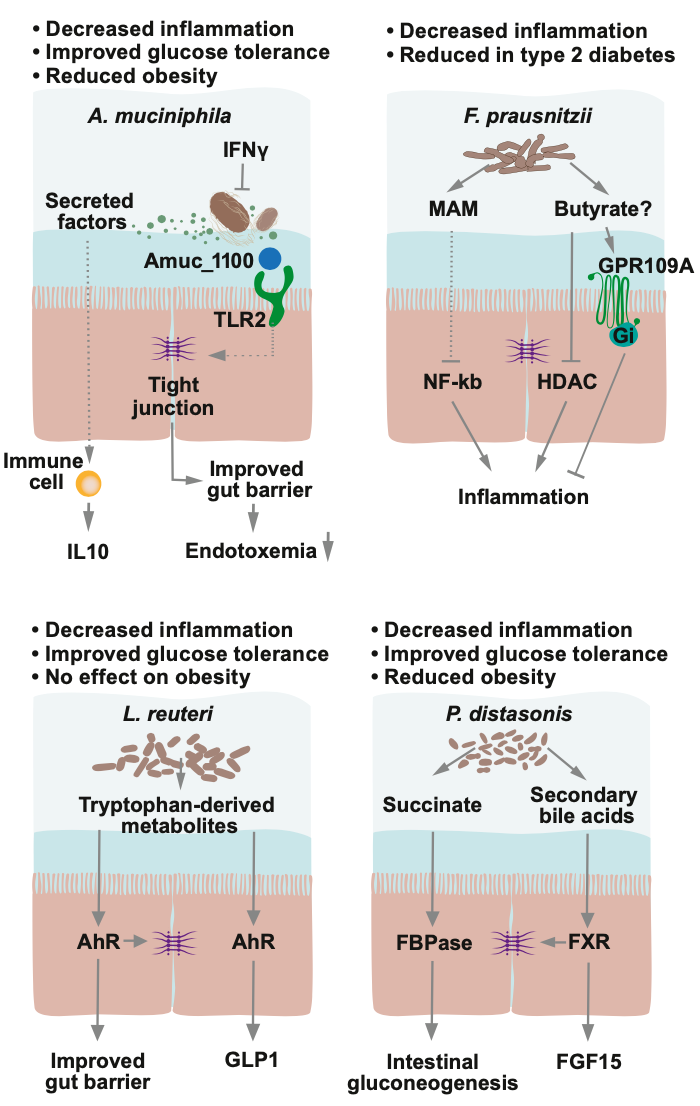
图 2. 有益细菌菌株及其作用机制
微生物代谢物:因果中的关键
肠道微生物可以产生众多的微生物代谢物,比如众所周知的短链脂肪酸(SCFAs)、胆汁酸和三甲胺 N-氧化物(TMAO),那么这些代谢物对宿主都有什么作用呢?
短链脂肪酸
短链脂肪酸是微生物发酵膳食纤维的产物,包括乙酸、丙酸和丁酸等。这些短链脂肪酸可以通过激活 G 蛋白偶联受体(GPCRs)、抑制组蛋白去乙酰化酶(HDACs)或者作为能量物质对宿主健康产生影响24。
丁酸可以通过调节肠上皮细胞的 IL-18 分泌,从而抑制调控 T 细胞和生成 IL-10 的细胞。此外某些丁酸还能作用于 GPR109A 或者 HDAC,从而具有有抗炎作用24。
丁酸还可以激活在结肠细胞中的过氧化物酶增殖物激活受体 γ (PPARγ),从而诱导丁酸的 β-氧化消耗氧气,调控肠道氧含量32。除了丁酸,丙酸也可以激活 PPARγ33。因此短链脂肪酸对维持无氧环境和抑制耐氧致病菌的大量生长有重要作用(图 3)。
胆汁酸
储存在胆囊中的胆汁酸由肝脏中的胆固醇产生。当人们进餐时,会分泌胆汁酸帮助吸收饮食中的脂质。初级胆汁酸经常与牛磺酸或者甘氨酸结合并且在回肠中会被再吸收。
细菌能够表达分解胆汁酸的胆汁盐水解酶,从而阻止胆汁酸在小肠中的吸收。未与牛磺酸或者甘氨酸结合的胆汁酸到达结肠时,它们会被肠道微生物进一步转化成为次级胆汁酸,次级胆汁酸的种类很多,其中最常见的是石胆酸和脱氧胆酸。
肠道菌闪烁梭菌(Clostridium scindens),可以产生次级胆汁酸,与艰难梭菌(C. difficile)感染的抑制相关34。另外,胆汁酸可以通过核法尼样 X 受体(FXR)和武田 G 蛋白偶联受体 5(TGR5,也称为 Gpbar1)传递信号,在控制宿主代谢方面具有一定潜能35。
氧化三甲胺
微生物代谢胆碱、卵磷脂、甜菜碱和左旋肉碱产生三甲胺(TMA)36。接着,三甲胺在肝脏中被黄素单加氧酶 3 氧化成为氧化三甲胺(TMAO)。TMAO 与心血管代谢疾病比如动脉粥样硬化、二型糖尿病、心肌梗塞等有关37,38。
关于 TMAO 的一个矛盾之处在于:鱼类和海鲜含有一定量的 TMAO,但是通常来讲这类食物具有保护心血管作用30。对于该矛盾,一个可能的解释是微生物的 TMA 产生会导致胆碱不足,从而通过影响 DNA 甲基化造成代谢疾病恶化,然而,通过饮食摄入的 TMAO 可能不会引起胆碱的不足或者可能不具有生物活性40。
此外另一个矛盾是,当高脂肪饮食小鼠规律摄入 TMAO 时,会造成小鼠的葡萄糖耐受性恶化,但是皮下注射 TMAO 却可以改善葡萄糖动态平衡和胰岛素分泌41,42。因此,仍需对 TMAO 的作用进一步研究。
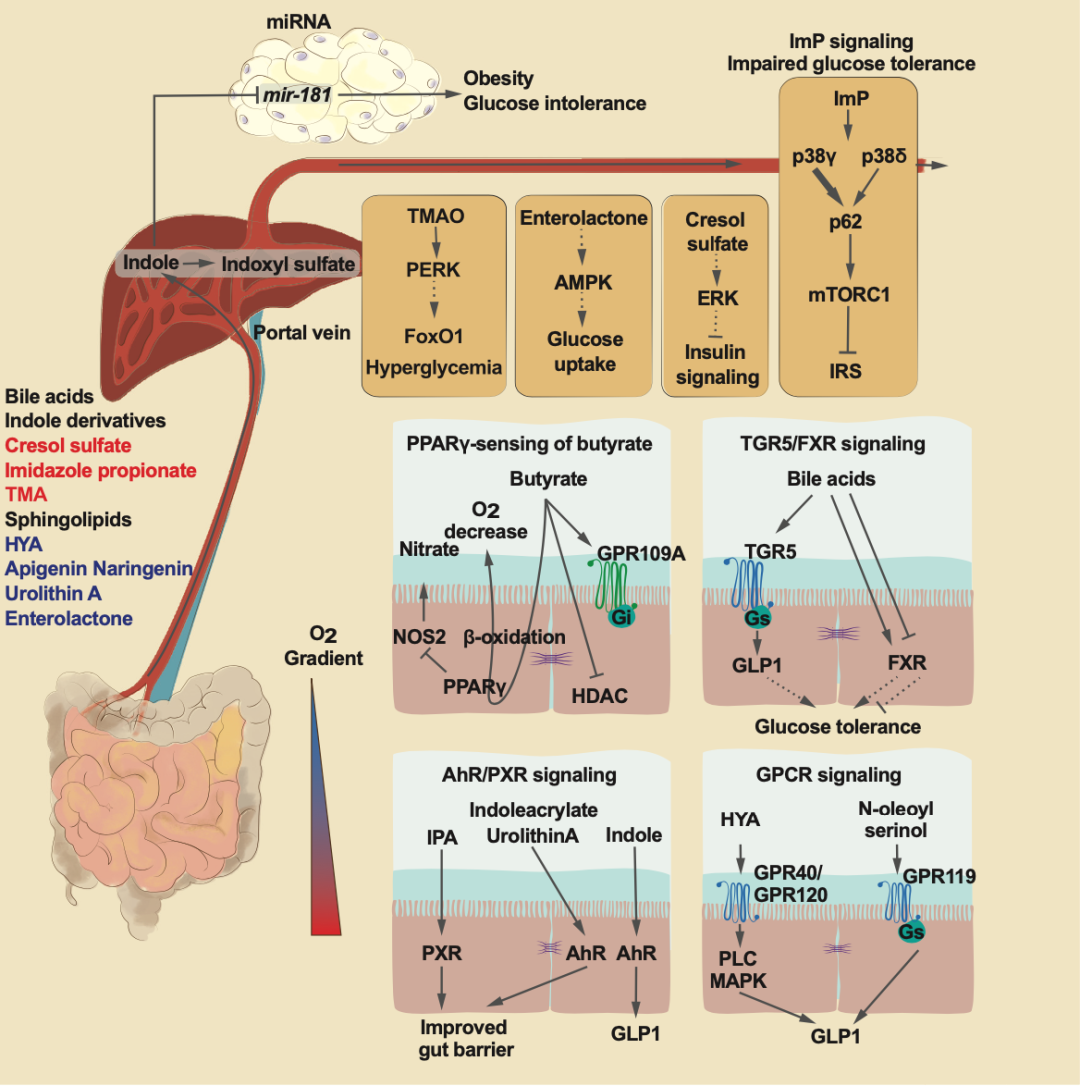
图 3. 微生物代谢物影响健康和疾病的机制
色氨酸衍生物
革兰氏阳性细菌和革兰氏阴性细菌中有一些微生物可以将色氨酸代谢产生多种产物(图 4),包括吲哚、吲哚乙酸、色胺、吲哚醛、吲哚乳酸和吲哚丙烯酸酯,其中部分衍生物可以激活受体 AhR7,43~45。
在小鼠中,研究发现 AhR 的激活可以有效减轻炎症,维持肠道动态平衡44,45。然而,AhR 在肝脏中的过量表达和非微生物 AhR 激动剂 TCDD(2,3,7,8-tetrachlorodibenzo-p-dioxin),会诱导正常饲料喂养小鼠发生肝脂肪变性46。
因此 AhR 激活不是绝对有益的,可能有环境和组织特异性效应。另外局部浓度是否足够激活目标受体也同样重要。
另外,研究还发现 AhR 的配体响应能力是具有种物种特异性的(表 2):TCDD 对鼠的亲和力是人类 AhR 的 10 倍;吲哚只对人类 AhR 起作用;硫酸吲哚酚对人体 AhR 的作用比鼠中高 500 倍48,49。
此外,在人体中,通过 TCDD 延长 AhR 的活性与毒性反应相关,因此,通过微生物产生的色氨酸衍生物引发的 AhR 活性可能在人体中会产生毒性(与在小鼠中的报道相反)。
目前为止,唯一的特例是吲哚丙烯酸,它是唯一一种在人体和鼠衍生的细胞系中作用没有差异的物质。
吲哚丙酸(IPA)不是 AhR 的配体,它与降低二型糖尿病的风险相关,口服 IPA 的大鼠表现出葡萄糖代谢的改善50,51。IPA 还会引起鼠中的抗炎症效果,但是在人体细胞中没有产生这种效果45。
除了 AhR 以外,小 RNA mir-181 被证实也是吲哚和其代谢物硫酸吲哚酚的靶标,因为这些代谢物表现出对 mir-181 表达的抑制。在高脂肪饮食小鼠中,吲哚能够有效预防肥胖47。而且,研究发现,至少在小鼠中,吲哚的这种益处与 AhR 和 mir-181 有关,但是在人体中尚不明确。
总之,上述结论提示我们在将色氨酸衍生物用于治疗之前,需要进一步阐明它在人体中的效果。
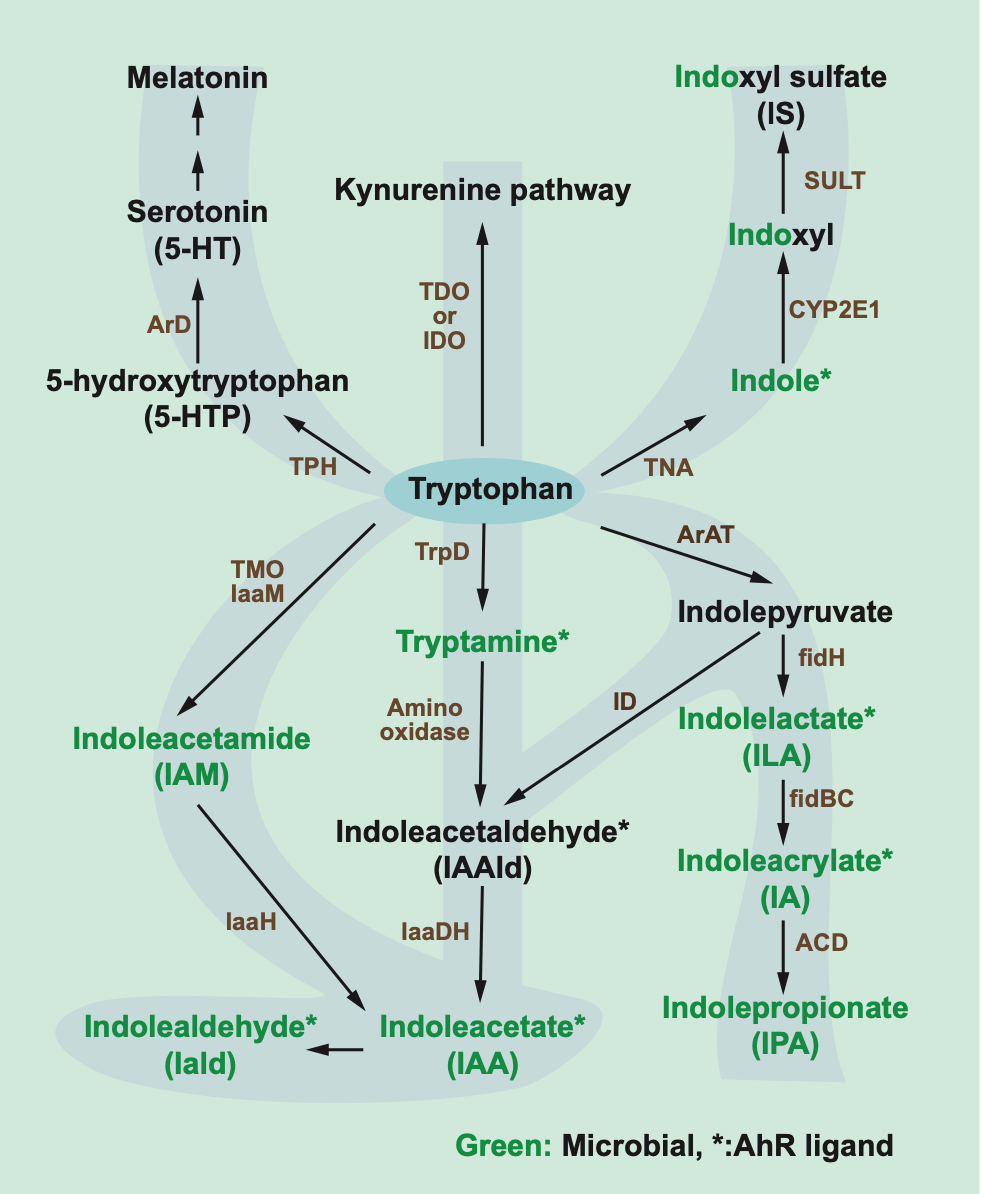
图4. 色氨酸降解途径
酪氨酸衍生物
酪氨酸能够被肠道微生物分解为对甲酚52。对甲酚在结肠粘膜和肝脏中会被进一步转化为对甲酚硫酸盐。对甲酚硫酸盐和吲哚氧基硫酸盐的提高与 CKD 死亡率增加相关,并且对甲酚硫酸盐(但不包括吲哚氧基硫酸盐)与心血管事件风险的增加也有关53。
致病菌艰难梭菌中产生的对甲酚赋予了这种致病菌优于其他共生菌的生存优势(尤其是革兰氏阴性菌)54。因此,将细菌的对甲酚代谢途径作为目标或可改变微生物组成并改善肠道健康。
组氨酸衍生物
在细菌和人体中,传统的组氨酸降解会产生尿刊酸和谷氨酸。大约 50 年前,尿刊酸代谢产物咪唑丙酸就被认为是肠道不健康的标志。最近,研究发现了多种含有尿刊酸还原酶(UrdA)的细菌,并且发现在一些拟杆菌、疣微菌和产丁酸细菌中不存在 UrdA55。由于尿刊酸是一种电子受体,所以在电子受体很少的环境中,可能含 UrdA 的细菌会更加丰富。
最近的研究发现咪唑丙酸在健康方面存在有害作用。首先,在二型糖尿病的样品中,咪唑丙酸的血浆浓度比在正常葡萄糖耐受的对照中高55。其次,咪唑丙酸与微生物多样性呈负相关56。第三,咪唑丙酸可以诱导小鼠中的葡萄糖不耐受,并且损害 IRS 介导的胰岛素信号55(图 3)。
咪唑丙酸的作用目前并没有表现出物种特异性,它在小鼠体内、鼠衍生的初级干细胞、人类胚胎肾细胞中均有类似作用(表 2)。
具有生物活性的脂质
肠道微生物也可能影响脂质的代谢,通过提高多不饱和脂肪酸的饱和度来产生一系列的脂肪酸(如共轭脂肪酸、羟基脂肪酸和含氧脂肪酸)57。10-羟基-顺式-12-十八碳稀酸(HYA),是由微生物代谢产生的初始中间共轭亚油酸,发现具有阻止高脂饮食诱导的肥胖和葡萄糖不耐受作用58。
鞘脂类物质是结构膜的成分,也是真核生物中重要的信号分子。宿主产生的鞘磷脂比如神经酰胺、鞘磷脂、鞘氨醇,在其鞘氨基醇骨架上存在一个双键(4,5-反式双键)59。抑制这种双键的插入(导致饱和二氢神经酰胺的增多)可以解决小鼠肝脏脂肪变性和胰岛素抵抗60。
有意思的是,细菌产的神经酰胺在鞘氨基醇骨架上不会有双键,因此,细菌的二氢神经酰胺水平是否在胰岛素敏感样品中较高以及是否会影响胰岛素信号,将会是一个有趣的研究。
拟杆菌衍生的鞘脂类对于维持肠道稳态具有关键作用59。脱氧鞘脂的 C1 位缺少一个羟基,不能被进一步代谢,因此一旦产生会被积累。脱氧鞘脂是胰岛 β 细胞的细胞毒性物质,并且血浆脱氧鞘磷脂水平已经成为 2 型糖尿病的风险预测物61,62。然而,微生物产生的脱氧鞘磷脂对糖尿病的具体作用还没有研究。
此外,还发现微生物产生的 N-oleoyl serinol 具有 GPCR 活性,而 GPCR 与胰岛素分泌有关。因此该物质也有可能是一个新的治疗靶点,但需要首先确定的是,这种代谢物的水平是否会因糖尿病状态的不同而不同。
多酚类物质
多酚类化合物可分为黄酮类化合物(黄酮醇、异黄酮类化合物、黄烷-3-醇类化合物、花青素-二丁烷类化合物)、酚酸类化合物、木脂素类化合物和芪类化合物(白藜芦醇)。
大部分膳食多酚类被微生物酶去糖基化、脱羟基化、去甲基化、减少碳碳双键并降解63~66。因此,微生物对黄酮类物质的降解作用可能降低富含黄酮类饮食的营养价值67。
但是微生物也能够通过产生多酚类衍生物,比如尿石素 A 和肠内脂,发挥潜在的益处。尿石素 A 是一种微生物衍生的代谢物,从天然多酚鞣花酸通过对一个内酯环脱羧并连续清除羟基形成64,66。
研究发现,尿石素 A 可以通过 AhR 激活增强小鼠中的肠道屏障(图3),但是在一些人类细胞系中它是 AhR 的拮抗剂,抑制吲哚或者硫酸吲哚诱导的人体 AhR 活性(表 2)68,69。最近一项关于尿石素 A 的临床试验表明,补充尿石素 A 老年人的线粒体和细胞健康状况得到改善,并且安全性良好70。
在亚麻籽中含量较多的木脂素能够被肠道微生物代谢成肠二醇,再被氧化成肠内酯,并进入循环中63,65。研究发现,在美国女性中,微生物肠内脂水平与 2 型糖尿病的低风险相关。此外,在小鼠实验中发现,肠内脂可以提高遗传性糖尿病小鼠的葡萄糖耐受71,72。
总的来说,多酚类物质依赖于微生物的酶活性,并且可能具有不同的代谢效果,了解细菌代谢多酚的途径可能较为重要。
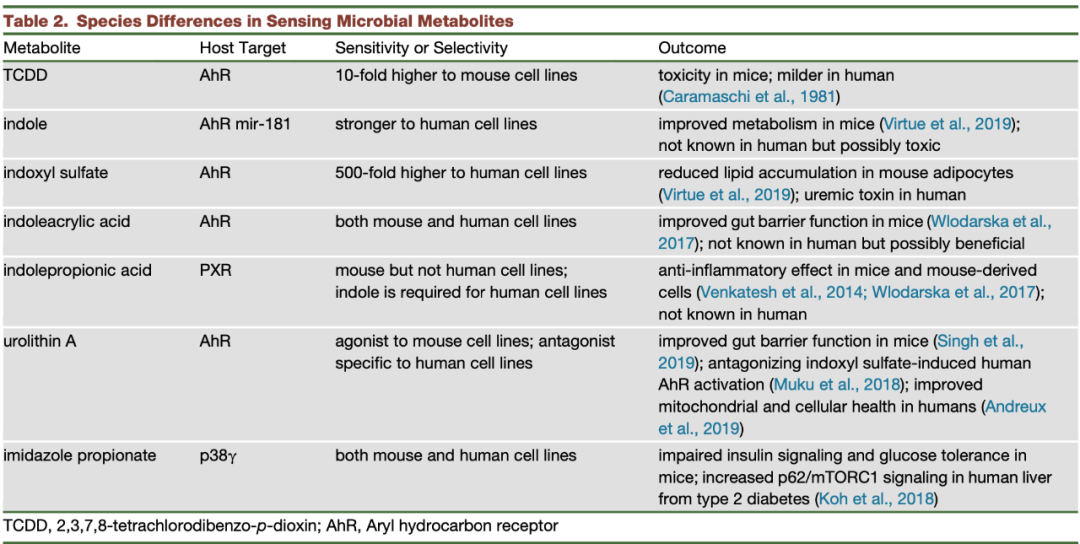
表 2 微生物代谢物在不同物种中的感知差异
未来的方向
尽管目前的技术得到了很大的发展,但是微生物组中大约 50%的基因还是未知的,此外由于注释方法的局限,导致在微生物基因、酶、产物、代谢物之间存在鸿沟。因此,联合多种计算方法对多组学数据的分析是很关键的73。
单一的代谢物测量可能不能提供准确的信息,因为已经有关于微生物代谢组昼夜节律的报道74。鉴于微生物改变后代谢组的变化具有一定的延迟性,因此在进行关联研究时,必须谨慎。此外,在确定饮食、微生物和宿主之间的精确和动态相互作用时需要记录多天饮食、纵向抽样(或时间系列取样)并和同位素标记代谢组学、宏基因组学等多种信息结合。
人体和鼠响应代谢物之间的差别是需要重视的,特别当把在小鼠中的试验结果转移到人身上时尤其要慎重。因此,先对人体细胞系进行实验是很重要的。
另外,代谢物并不会单独起作用,因此需要研究不同代谢物介导的联合信号。针对这一问题,可以在先前确定的微生物代谢物目标之外,确定更多的目标,以更好地捕获单个代谢物的全部信号能力。还可以通过建立一个具有明确微生物群落的定菌小鼠模型来研究特定微生物代谢产物对宿主代谢的作用40,75。
在治疗潜能方面,一些微生物菌株已经在人体中实验,但是它们对代谢的影响看起来并不大。另外,因为宿主的差异,益生菌可能表现出非预期的结果76,因此,鉴定益生菌的非活成分比如分泌物、多肽和膜成分介导的益生效果,可能会更加有用。
针对细菌成分制作疫苗是一种治疗代谢疾病的潜在方法,因此鉴定有效的细菌成分也是关键。开发针对细菌酶的抑制剂则是另一种预防有害微生物代谢物的策略。
综合来讲,微生物组为我们提供了一系列有吸引力的手段——改变个性化饮食、创造特定的下一代益生菌、针对微生物的酶,这些方法都可以用来改变人类的代谢,治疗代谢疾病。
参考文献:
(滑动下方文字查看)
1.Sender,R., Fuchs, S., and Milo, R. (2016). Are We Really Vastly Outnumbered?Revisiting the Ratio of Bacterial to Host Cells in Humans. Cell 164, 337–340.
2.Tierney,B.T., Yang, Z., Luber, J.M., Beaudin, M., Wibowo, M.C., Baek, C.,Mehlenbacher,E., Patel, C.J., and Kostic, A.D. (2019). The Landscape of Genetic Content inthe Gut and Oral Human Microbiome. Cell Host Microbe 26, 283–295.
3.Maier,L., Pruteanu, M., Kuhn, M., Zeller, G., Telzerow, A., Anderson, E.E., Brochado,A.R., Fernandez, K.C., Dose, H., Mori, H., et al. (2018). Extensive impact ofnon-antibiotic drugs on human gut bacteria. Nature 555, 623–628.
4.Sonnenburg,J.L., and Backhed, F. (2016). Diet-microbiota interactions as moderators ofhuman metabolism. Nature 535, 56–64.
5.Sun,L.L., Xie, C., Wang, G., Wu, Y., Wu, Q., Wang, X.M., Liu, J., Deng, Y.Y., Xia,J.L., Chen, B., et al. (2018). Gut microbiota and intestinal FXR mediate theclinical benefits of metformin. Nat. Med. 24, 1919.
6.Wu,H., Esteve, E., Tremaroli, V., Khan, M.T., Caesar, R., Manneras-Holm, L., Stahlman,M., Olsson, L.M., Serino, M., Planas-Felix, M., et al. (2017). Metformin altersthe gut microbiome of individuals with treatment-naive type 2 diabetes,contributing to the therapeutic effects of the drug. Nat. Med. 23, 850.
7.Natividad,J.M., Agus, A., Planchais, J., Lamas, B., Jarry, A.C., Martin, R., Michel,M.L., Chong-Nguyen, C., Roussel, R., Straube, M., et al. (2018). Impaired ArylHydrocarbon Receptor Ligand Production by the Gut Microbiota Is a Key Factor inMetabolic Syndrome. Cell Metab. 28, 737–749.
8.Roager,H.M., Vogt, J.K., Kristensen, M., Hansen, L.B.S., Ibrugger, S., Maerkedahl,R.B., Bahl, M.L., Lind, M.V., Nielsen, R.L., Frokiaer, H., et al. (2019). Wholegrain-rich diet reduces body weight and systemic low-grade inflammation withoutinducing major changes of the gut microbiome: a randomized cross-over trial.Gut 68, 83–93.
9.Zhao,L.P., Zhang, F., Ding, X.Y., Wu, G.J., Lam, Y.Y., Wang, X.J., Fu, H.Q., Xue,X.H., Lu, C.H., Ma, J.L., et al. (2018). Gut bacteria selectively promoted bydietary fibers alleviate type 2 diabetes. Science 359, 1151–1156.
10.Uchimura,Y., Fuhrer, T., Li, H., Lawson, M.A., Zimmermann, M., Yilmaz, B., Zindel, J.,Ronchi, F., Sorribas, M., Hapfelmeier, S., et al. (2018). Antibodies SetBoundaries Limiting Microbial Metabolite Penetration and the Resultant MammalianHost Response. Immunity 49, 545–559.
11.Khan,M.T., Nieuwdorp, M., and Backhed, F. (2014). Microbial Modulation of Insulin Sensitivity.Cell Metab. 20, 753–760.
12.Backhed,F., Manchester, J.K., Semenkovich, C.F., and Gordon, J.I. (2007). Mechanismsunderlying the resistance to diet-induced obesity in germ-free mice. Proc.Natl. Acad. Sci. USA 104, 979–984.
13.Turnbaugh,P.J., Ley, R.E., Mahowald, M.A., Magrini, V., Mardis, E.R., and Gordon, J.I.(2006). An obesity-associated gut microbiome with increased capacity for energyharvest. Nature 444, 1027–1031.
14.Ridaura,V.K., Faith, J.J., Rey, F.E., Cheng, J.Y., Duncan, A.E., Kau, A.L., Griffin,N.W., Lombard, V., Henrissat, B., Bain, J.R., et al. (2013). Gut Microbiotafrom Twins Discordant for Obesity Modulate Metabolism in Mice. Science 341, https://doi.org/10.1126/science.1241214.
15.vanNood, E., Vrieze, A., Nieuwdorp, M., Fuentes, S., Zoetendal, E.G., de Vos, W.M.,Visser, C.E., Kuijper, E.J., Bartelsman, J.F.W.M., Tijssen, J.G.P., et al. (2013).Duodenal Infusion of Donor Feces for Recurrent Clostridium difficile. N. Engl.J. Med. 368, 407–415.
16.Alang,N., and Kelly, C.R. (2015). Weight Gain After Fecal Microbiota Transplantation.Open Forum Infect Di 2 2, https://doi.org/10.1093/ofid/ofv004.
17.Vrieze,A., Van Nood, E., Holleman, F., Salojarvi, J., Kootte, R.S., Bartelsman, J.F.W.M.,Dallinga-Thie, G.M., Ackermans, M.T., Serlie, M.J., Oozeer, R., et al. (2012).Transfer of Intestinal Microbiota From Lean Donors Increases Insulin Sensitivityin Individuals With Metabolic Syndrome. Gastroenterology 143, 913–916.
18.DeFilipp,Z., Bloom, P.P., Torres Soto, M., Mansour, M.K., Sater, M.R.A., Huntley, M.H.,Turbett, S., Chung, R.T., Chen, Y.-B., and Hohmann, E.L. (2019). Drug-ResistantE. coli Bacteremia Transmitted by Fecal Microbiota Transplant. N. Engl. J. Med.381, 2043–2050.
19.Derrien,M., Collado, M.C., Ben-Amor, K., Salminen, S., and de Vos, W.M. (2008). Themucin degrader Akkermansia muciniphila is an abundant resident of the humanintestinal tract. Appl. Environ. Microbiol. 74, 1646–1648.
20.Everard,A., Belzer, C., Geurts, L., Ouwerkerk, J.P., Druart, C., Bindels, L.B., Guiot,Y., Derrien, M., Muccioli, G.G., Delzenne, N.M., et al. (2013). Crosstalkbetween Akkermansia muciniphila and intestinal epithelium controls diet-inducedobesity. Proc. Natl. Acad. Sci. USA 110, 9066–9071.
21.Depommier,C., Everard, A., Druart, C., Plovier, H., Van Hul, M., Vieira-Silva, S., Falony,G., Raes, J., Maiter, D., Delzenne, N.M., et al. (2019). Supplementation withAkkermansia muciniphila in overweight and obese human volunteers: a proof-of-conceptexploratory study. Nat. Med. 25, 1096–1103.
22.Ottman,N., Reunanen, J., Meijerink, M., Pietila, T.E., Kainulainen, V., Klievink, J.,Huuskonen, L., Aalvink, S., Skurnik, M., Boeren, S., et al. (2017). Pili-like proteinsof Akkermansia muciniphila modulate host immune responses and gut barrierfunction. PLoS ONE 12, e0175766.
23.Plovier,H., Everard, A., Druart, C., Depommier, C., Van Hul, M., Geurts, L., Chilloux,J., Ottman, N., Duparc, T., Lichtenstein, L., et al. (2017). A purified membraneprotein from Akkermansia muciniphila or the pasteurized bacterium improvesmetabolism in obese and diabetic mice. Nat. Med. 23, 107–113.
24.Koh,A., De Vadder, F., Kovatcheva-Datchary, P., and Backhed, F. (2016). FromDietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key BacterialMetabolites. Cell 165, 1332–1345.
25.Quevrain,E., Maubert, M.A., Michon, C., Chain, F., Marquant, R., Tailhades, J., Miquel,S., Carlier, L., Bermudez-Humaran, L.G., Pigneur, B., et al. (2016). Identificationof an anti-inflammatory protein from Faecalibacterium prausnitzii, a commensalbacterium deficient in Crohn’s disease. Gut 65, 415–425.
26.Hippe,B., Remely, M., Aumueller, E., Pointner, A., Magnet, U., and Haslberger, A.G.(2016). Faecalibacterium prausnitzii phylotypes in type two diabetic, obese,and lean control subjects. Benef. Microbes 7, 511–517.
27.Udayappan,S., Manneras-Holm, L., Chaplin-Scott, A., Belzer, C., Herrema, H.,Dallinga-Thie, G.M., Duncan, S.H., Stroes, E.S.G., Groen, A.K., Flint, H.J., etal. (2016). Oral treatment with Eubacterium hallii improves insulin sensitivityin db/db mice. Npj Biofilms Microbi 2.
28.Wang,K., Liao, M.F., Zhou, N., Bao, L., Ma, K., Zheng, Z.Y., Wang, Y.J., Liu, C., Wang,W.Z., Wang, J., et al. (2019). Parabacteroides distasonis Alleviates Obesityand Metabolic Dysfunctions via Production of Succinate and Secondary BileAcids. Cell Rep. 26, 222–235.
29.DeVadder, F., Kovatcheva-Datchary, P., Zitoun, C., Duchampt, A., Backhed, F., andMithieux, G. (2016). Microbiota-Produced Succinate Improves Glucose Homeostasisvia Intestinal Gluconeogenesis. Cell Metab. 24, 151–157.
30.Kovatcheva-Datchary,P., Nilsson, A., Akrami, R., Lee, Y.S., De Vadder, F., Arora, T., Hallen, A.,Martens, E., Bjorck, I., and Backhed, F. (2015). Dietary FiberInducedImprovement in Glucose Metabolism Is Associated with Increased Abundance ofPrevotella. Cell Metab. 22, 971–982.
31.Wu,T.R., Lin, C.S., Chang, C.J., Lin, T.L., Martel, J., Ko, Y.F., Ojcius, D.M.,Lu, C.C., Young, J.D., and Lai, H.C. (2019). Gut commensal Parabacteroidesgoldsteinii plays a predominant role in the anti-obesity effects ofpolysaccharides isolated from Hirsutella sinensis. Gut 68, 248–262.
32.Byndloss,M.X., Olsan, E.E., Rivera-Chavez, F., Tiffany, C.R., Cevallos, S.A., Lokken,K.L., Torres, T.P., Byndloss, A.J., Faber, F., Gao, Y.D., et al. (2017). Microbiota-activatedPPAR-gamma signaling inhibits dysbiotic Enterobacteriaceae expansion. Science357, 570–575.
33.Nepelska,M., de Wouters, T., Jacouton, E., Beguet-Crespel, F., Lapaque, N., Dore, J.,Arulampalam, V., and Blottiere, H.M. (2017). Commensal gut bacteria modulatephosphorylation-dependent PPAR gamma transcriptional activity in humanintestinal epithelial cells. Sci Rep 7, https://doi.org/10.1038/ srep43199.
34.Buffie,C.G., Bucci, V., Stein, R.R., McKenney, P.T., Ling, L.L., Gobourne, A., No, D.,Liu, H., Kinnebrew, M., Viale, A., et al. (2015). Precision microbiome reconstitutionrestores bile acid mediated resistance to Clostridium difficile. Nature 517,205–208.
35.Wahlstrom,A., Sayin, S.I., Marschall, H.U., and Backhed, F. (2016). Intestinal Crosstalkbetween Bile Acids and Microbiota and Its Impact on Host Metabolism. CellMetab. 24, 41–50.
36.Koeth,R.A., Lam-Galvez, B.R., Kirsop, J., Wang, Z.N., Levison, B.S., Gu, X.D., Copeland,M.F., Bartlett, D., Cody, D.B., Dai, H.J., et al. (2019). L-Carnitineinomnivorous diets induces an atherogenic gut microbial pathway in humans. J.Clin. Invest. 129, 373–387.
37.Shan,Z.L., Sun, T.P., Huang, H., Chen, S.J., Chen, L.K., Luo, C., Yang, W., Yang,X.F., Yao, P., Cheng, J.Q., et al. (2017). Association betweenmicrobiota-dependent metabolite trimethylamine-N-oxide and type 2 diabetes. Am.J. Clin. Nutr. 106, 888–894.
38.Tang,W.H.W., Kitai, T., and Hazen, S.L. (2017). Gut Microbiota in CardiovascularHealth and Disease. Circ. Res. 120, 1183–1196.
39.Landfald,B., Valeur, J., Berstad, A., and Raa, J. (2017). Microbial trimethylamine-N-oxideas a disease marker: something fishy? Microb. Ecol. Health Dis. 28, 1327309.
40.Romano,K.A., Martinez-del Campo, A., Kasahara, K., Chittim, C.L., Vivas, E.I., Amador-Noguez,D., Balskus, E.P., and Rey, F.E. (2017). Metabolic, Epigenetic, andTransgenerational Effects of Gut Bacterial Choline Consumption.Cell HostMicrobe 22, 279–290.
41.Gao,X., Liu, X.F., Xu, J., Xue, C.H., Xue, Y., and Wang, Y.M. (2014). Dietary trimethylamineN-oxide exacerbates impaired glucose tolerance in mice fed a high fat diet. J.Biosci. Bioeng. 118, 476–481.
42.Dumas,M.E., Rothwell, A.R., Hoyles, L., Aranias, T., Chilloux, J., Calderari, S., Noll,E.M., Pean, N., Boulange, C.L., Blancher, C., et al. (2017). Microbial-Host Co-metabolitesAre Prodromal Markers Predicting Phenotypic Heterogeneity in Behavior, Obesity,and Impaired Glucose Tolerance. Cell Rep. 20, 136–148.
43.Zelante,T., Iannitti, R.G., Cunha, C., De Luca, A., Giovannini, G., Pieraccini, G., Zecchi,R., D’Angelo, C., Massi-Benedetti, C., Fallarino, F., et al. (2013). TryptophanCatabolites from Microbiota Engage Aryl Hydrocarbon Receptor and BalanceMucosal Reactivity via Interleukin-22. Immunity 39, 372–385.
44.Cervantes-Barragan,L., Chai, J.N., Tianero, M.D., Di Luccia, B., Ahern, P.P., Merriman, J.,Cortez, V.S., Caparon, M.G., Donia, M.S., Gilfillan, S., et al. (2017).Lactobacillus reuteri induces gut intraepithelial CD4(+)CD8 alpha alpha(+) Tcells. Science 357, 806–810.
45.Wlodarska,M., Luo, C.W., Kolde, R., d’Hennezel, E., Annand, J.W., Heim, C.E., Krastel,P., Schmitt, E.K., Omar, A.S., Creasey, E.A., et al. (2017). Indoleacrylic AcidProduced by Commensal Peptostreptococcus Species Suppresses Inflammation. CellHost Microbe 22, 25–37.
46.Lee,J.H., and Lee, J. (2010). Indole as an intercellular signal in microbial communities.FEMS Microbiol. Rev. 34, 426–444.
47.Virtue,A.T., McCright, S.J., Wright, J.M., Jimenez, M.T., Mowel, W.K., Kotzin, J.J.,Joannas, L., Basavappa, M.G., Spencer, S.P., Clark, M.L., et al. (2019). Thegut microbiota regulates white adipose tissue inflammation and obesity via afamily of microRNAs. Sci. Transl. Med. 11,https://doi.org/10.1126/scitranslmed.aav1892.
48.Hubbard,T.D., Murray, I.A., Bisson, W.H., Lahoti, T.S., Gowda, K., Amin, S.G., Patterson,A.D., and Perdew, G.H. (2015). Adaptation of the human arylhydrocarbon receptorto sense microbiota-derived indoles. Sci Rep. 5, https://doi.org/10.1038/srep12689.
49.Schroeder,J.C., DiNatale, B.C., Murray, I.A., Flaveny, C.A., Liu, Q., Laurenzana, E.M.,Lin, J.M., Strom, S.C., Omiecinski, C.J., Amin, S., et al. (2010). The UremicToxin 3-Indoxyl Sulfate Is a Potent Endogenous Agonist for the Human ArylHydrocarbon Receptor. Biochemistry 49, 393–400.
50.deMello, V.D., Paananen, J., Lindstrom, J., Lankinen, M.A., Shi, L., Kuusisto, J.,Pihlajamaki, J., Auriola, S., Lehtonen, M., Rolandsson, O., et al. (2017).Indolepropionic acid and novel lipid metabolites are associated with a lowerrisk of type 2 diabetes in the Finnish Diabetes Prevention Study. Sci Rep. https://doi.org/10.1038/srep46337.
51.Abildgaard,A., Elfving, B., Hokland, M., Wegener, G., and Lund, S. (2018). The microbialmetabolite indole-3-propionic acid improves glucose metabolism in rats, butdoes not affect behaviour. Arch. Physiol. Biochem. 124, 306–312.
52.Smith,E.A., and Macfarlane, G.T. (1996). Enumeration of human colonic bacteriaproducing phenolic and indolic compounds: Effects of pH, carbohydrate availabilityand retention time on dissimilatory aromatic amino acid metabolism. J. Appl. Bacteriol.81, 288–302.
53.Lin,C.J., Wu, V., Wu, P.C., and Wu, C.J. (2015). Meta-Analysis of the Associationsof p-Cresyl Sulfate (PCS) and Indoxyl Sulfate (IS) with Cardiovascular Eventsand All-Cause Mortality in Patients with Chronic Renal Failure. PLoS ONE 10,e0132589.
54.Passmore,I.J., Letertre, M.P.M., Preston, M.D., Bianconi, I., Harrison, M.A., Hasher,F., Kaur, H., Hong, H.A., Baines, S.D., Cutting, S.M., et al. (2018). Para-cresolproduction by Clostridium difficile affects microbial diversity and membraneintegrity of Gram-negative bacteria. PLoS Pathog. 14, e1007191.
55.Koh,A., Molinaro, A., Stahlman, M., Khan, M.T., Schmidt, C., Manneras-Holm, L., Wu,H., Carreras, A., Jeong, H., Olofsson, L.E., et al. (2018). Microbially ProducedImidazole Propionate Impairs Insulin Signaling through mTORC1. Cell 175,947–961.
56.Wilmanski,T., Rappaport, N., Earls, J.C., Magis, A.T., Manor, O., Lovejoy, J., Omenn,G.S., Hood, L., Gibbons, S.M., and Price, N.D. (2019). Blood metabolomepredicts gut microbiome a-diversity in humans. Nat. Biotechnol. 37, 1217–1228.
57.Kishino,S., Takeuchi, M., Park, S.B., Hirata, A., Kitamura, N., Kunisawa, J., Kiyono,H., Iwamoto, R., Isobe, Y., Arita, M., et al. (2013). Polyunsaturated fatty acidsaturation by gut lactic acid bacteria affecting host lipid composition. Proc.Natl. Acad. Sci. USA 110, 17808–17813.
58.Miyamoto,J., Igarashi, M., Watanabe, K., Karaki, S., Mukouyama, H., Kishino, S., Li, X.,Ichimura, A., Irie, J., Sugimoto, Y., et al. (2019). Gut microbiota confershost resistance to obesity by metabolizing dietary polyunsaturated fatty acids.Nat. Commun. 10,, https://doi.org/10.1038/s41467-019-11978-0.
59.Brown,E.M., Ke, X.B., Hitchcock, D., Jeanfavre, S., Avila-Pacheco, J., Nakata, T.,Arthur, T.D., Fornelos, N., Heim, C., Franzosa, E.A., et al. (2019). Bacteroides-DerivedSphingolipids Are Critical for Maintaining Intestinal Homeostasis andSymbiosis. Cell Host Microbe 25, 668–680.
60.Chaurasia,B., Tippetts, T.S., Monibas, R.M., Liu, J.Q., Li, Y., Wang, L.P., Wilkerson,J.L., Sweeney, C.R., Pereira, R.F., Sumida, D.H., et al. (2019). Targeting aceramide double bond improves insulin resistance and hepatic steatosis. Science365, 386–392.
61.Zuellig,R.A., Hornemann, T., Othman, A., Hehl, A.B., Bode, H., Guntert, T., Ogunshola,O.O., Saponara, E., Grabliauskaite, K., Jang, J.H., et al. (2014).Deoxysphingolipids, novel biomarkers for type 2 diabetes, are cytotoxic forinsulin-producing cells. Diabetes 63, 1326–1339.
62.Mwinyi,J., Bostrom, A., Fehrer, I., Othman, A., Waeber, G., Marti-Soler, H., Vollenweider,P., Marques-Vidal, P., Schioth, H.B., von Eckardstein, A., et al. (2017).Plasma 1-deoxysphingolipids are early predictors of incident type 2 diabetesmellitus (vol 12, e0175776, 2017). PLoS ONE 12, e0175776.
63.Borriello,S.P., Setchell, K.D., Axelson, M., and Lawson, A.M. (1985). Production andmetabolism of lignans by the human faecal flora. J. Appl. Bacteriol. 58, 37–43.
64.Cerda,B., Periago, P., Espin, J.C., and Tomas-Barberan, F.A. (2005). Identificationof urolithin A as a metabolite produced by human colon microflora from ellagicacid and related compounds. J. Agric. Food Chem. 53, 5571–5576.
65.Clavel,T., Henderson, G., Engst, W., Dore, J., and Blaut, M. (2006). Phylogeny ofhuman intestinal bacteria that activate the dietary lignan secoisolariciresinoldiglucoside. FEMS Microbiol. Ecol. 55, 471–478.
66.Espin,J.C., Larrosa, M., Garcia-Conesa, M.T., and Tomas-Barberan, F. (2013).Biological Significance of Urolithins, the Gut Microbial Ellagic AcidDerivedMetabolites: The Evidence So Far. Evid-Based Complement Alternate Med. 2013,270418.
67.Thaiss,C.A., Tav, S.I., Rothschild, D., Eijer, M.T.M., Levy, M., Moresi, C.,Dohnalova, L., Braverman, S., Rozin, S., Malitsky, S., et al. (2016b).Persistent microbiome alterations modulate the rate of post-dieting weightregain. Nature 540, 544–551.
68.Singh,R., Chandrashekharappa, S., Bodduluri, S.R., Baby, B.V., Hegde, B., Kotla,N.G., Hiwale, A.A., Saiyed, T., Patel, P., Vijay-Kumar, M., et al. (2019). Enhancementof the gut barrier integrity by a microbial metabolite through the Nrf2pathway. Nat. Commun. 10, https://doi.org/10.1038/s41467-018-07859-7.
69.Muku,G.E., Murray, I.A., Espin, J.C., and Perdew, G.H. (2018). Urolithin A Is a DietaryMicrobiota-Derived Human Aryl Hydrocarbon Receptor Antagonist. Metabolites 8, https://doi.org/10.3390/metabo8040086.
70.Andreux,P.A., Blanco-Bose, W., Ryu, D., Burdet, F., Ibberson, M., Aebischer, P.,Auwerx, J., Singh, A., and Rinsch, C. (2019). The mitophagy activator urolithinA is safe and induces a molecular signature of improved mitochondrial andcellular health in humans. Nature Metabolism 1, 595–603.
71.Sun,Q., Wedick, N.M., Pan, A., Townsend, M.K., Cassidy, A., Franke, A.A., Rimm,E.B., Hu, F.B., and van Dam, R.M. (2014). Gut Microbiota Metabolites of DietaryLignans and Risk of Type 2 Diabetes: A Prospective Investigation in Two Cohortsof U. S. Women. Diabetes Care 37, 1287–1295.
72.Zhou,F., Furuhashi, K., Son, M.J., Toyozaki, M., Yoshizawa, F., Miura, Y., and Yagasaki,K. (2017). Antidiabetic effect of enterolactone in cultured muscle cells and intype 2 diabetic model db/db mice. Cytotechnology 69, 493–502.
73.Joice,R., Yasuda, K., Shafquat, A., Morgan, X.C., and Huttenhower, C. (2014). DeterminingMicrobial Products and Identifying Molecular Targets in the Human Microbiome.Cell Metab. 20, 731–741.
74.Thaiss,C.A., Levy, M., Korem, T., Dohnalova, L., Shapiro, H., Jaitin, D.A., David, E.,Winter, D.R., Gury-BenAri, M., Tatirovsky, E., et al. (2016a). Microbiota DiurnalRhythmicity Programs Host Transcriptome Oscillations. Cell 167, 1495–1510e1412.
75.Kasahara,K., Krautkramer, K.A., Org, E., Romano, K.A., Kerby, R.L., Vivas, E.I.,Mehrabian, M., Denu, J.M., Backheds, F., Lusis, A., et al. (2018). Interactionsbetween Roseburia intestinalis and diet modulate atherogenesis in a murinemodel. Nat. Microbiol. 3, 1461–1471.
76.Suez,J., Zmora, N., Zilberman-Schapira, G., Mor, U., Dori-Bachash, M., Bashiardes,S., Zur, M., Regev-Lehavi, D., Brik, R.B., Federici, S., et al. (2018). Post-AntibioticGut Mucosal Microbiome Reconstitution Is Impaired by Probiotics and Improved byAutologous FMT. Cell 174, 1406–1423.
作者|gemiu
审校|617
编辑|笑咲





投稿/转载
联系人:何隽
微信号:18518006142